![]()
Research Interests |
||||||
Protein N-glycosylation and Golgi remodeling are essential in vertebrates.The cell membrane is a protective bilayer embedded with glycoproteins that mediate interactions interaction with the environment, such as cell-cell signaling and nutrient uptake. In the secretory pathway, newly synthesized proteins are co-translational modified by N-glycosylation at Asn-X-Ser/Thr (X≠Pro) sites [NXS/T] sites, and ancient modification that promotes folding and proteostasis in the ER. Glycoproteins transit from the ER to the Golgi in metazoans where the N-glycans are trimmed by glycosidases and modified by branching N-acetylglucosaminyltransferases (MGAT-1,-2,-3,-4, and -5). |
||||||
Computational model of N-glycan branching and opposing signaling pathways.
|
||||||
Genetic code asymmetry supports diversity through experimentation with posttranslational modifications.The topology of the genetic code clusters synonymous codons, and amino acids with similar chemical properties, which minimizes the deleterious effects of mutations and translation error. The evolutionary |
||||||
Essential amino acid exchangers are regulated by glycosylated adaptors (28).
|
||||||
Gene purging and the evolution of Neoave metabolism and longevity (29).Maintenance of the proteasome requires oxidative phosphorylation (ATP) and mitigation of oxidative damage, in an increasingly dysfunctional relationship with aging. SLC3A2 plays a role on both sides of this dichotomy as an adaptor to SLC7A5, a transporter of branched-chain amino acids (BCAA: Leu, Ile, Val), and to SLC7A11, a cystine importer supplying cysteine to the synthesis of the antioxidant glutathione. Intriguingly, Neoaves have purged CDKN1A (P21) and CDKN2A (P16) as well as two associated kinases CDK2 and CDK4 that play a role in metabolism, cell cycle and senescence. Further analysis of Neoave genomes may reveal higher-dimensional synthetic gene interactions beyond what can be achieved with inbred strains of mice and dogs. The evolution of birds may be viewed as a gain-of-function, gene drop-out experiment driven by adaptation to a volant lifestyle, which has resulted in a remarkable radiation of species occupying ecosystems worldwide and many with extended longevity. Further molecular analysis of Neoaves and Bats may reveal novel rewiring with applications for human health and longevity (29). |
||||||
Hexosamine biosynthesis pathway in C. elegans development, stress and aging.Dr. Wendy Johnston in our lab has shown that C. elegans require a large increase in UDP-GlcNAc very early in development for the synthesis of eggshell chitin, a GlcNAcβ1-4 polymer. Chitin forms a protective barrier that blocks entry by supernumerary sperm during fertilization. Synthesis of chitin depends on glucosamine-6-phosphate N-acetyltransferase 2 (GNA-2), a hexosamine pathway enzyme which is rate limiting for UDP-GlcNAc biosynthesis in the C. elegans early zygote (30-32). Gna-2 null mutants are maternal effect embryonic lethal. Rapid and asymmetric extrusion of eggshell chitin is required for movement of the sperm pronucleus to the cortex, as well as reorganization of the cytoskeleton and anterior-posterior polarization. Chitin is also crucial for high fidelity meiosis, polar body extrusion, and turnover of oocyte proteins during the cellular switch to a zygotic phenotype. A screen for gna-2 suppressors yielded mutants of sup-46 encoding an RNA-binding protein (C25A1.4) that localizes to stress granules and increases the gna-1 expression (33). Stress granules occur by phase transition at critical concentrations of interacting RNA and proteins (image below). Dr. Joe Culotti has identified genetic and environmental factors that act through protein glycosylation to navigate gonad distal tip cell (DTC) migration during embryogenesis (34). Working with Joe, we find that mutation of glycosyltransferases encoding branching in the N-glycosylation pathway, and elongation in the chondroitin pathway display defects in phase 2 (ventral to dorsal) migration of the DTC, with penetrance that is temperature sensitive. N-glycosylation and chondroitin mutations displayed additive penetrance (>90%), suggesting compensation and redundancy of these pathways in phase 2 DTC guidance. Both N-glycans and chondroitin have beta-galactoside sequences with affinity for Galectins (in preparation, 2024). Dr. Charles Warren, a talented postdoctoral fellow, initiated the C. elegans work in my lab. He was first to clone gly-2, which encoded worm MGAT5 activity and showed gly-2 can rescue an Mgat5 deficiency in mammalian cells (35,36). Working with Aldis Krizus, he also mapped the expression patterns six nonessential C. elegans core 2/I N-acetylglucosaminyltransferase homologues (37,38). Charles became assistant Professor at University of New Hampshire in 2003. He designed a study of congenital disorder of glycosylation in C. elegans; a genome-wide RNAi screen to identify N-glycosylation-dependent loci (39). Tragically, Charles died in a paragliding accident in 2005. |
||||||
Polo-like kinase-4Polo-like kinase 4 (Plk4) is a serine/threonine kinase first cloned in our Lab as named SAK, a homolog of polo kinase (PLK) and subsequently named PLK4 (40-42). Plk4 is required for centrosome duplication and fidelity of chromosome segregation at mitosis. Plk4 haploinsufficiency promotes mitotic instability and carcinogenesis (43,44). Work on Plk4 continues, led by Dr. Carol Swallow at LTRI. PLK4 is found at high levels in some aggressive cancers of the colorectum, pancreas and breast (45). Plk4 also regulates cancer cell migration and invasion (44,46,47). Plk4 inhibitors are being developed by Phara, and clinical trials in cancer patients are showing promise. |
||||||
| ► Pubmed — search for selected publications | ||||||
|
REFERENCES
|
||||||
|
|
||||||


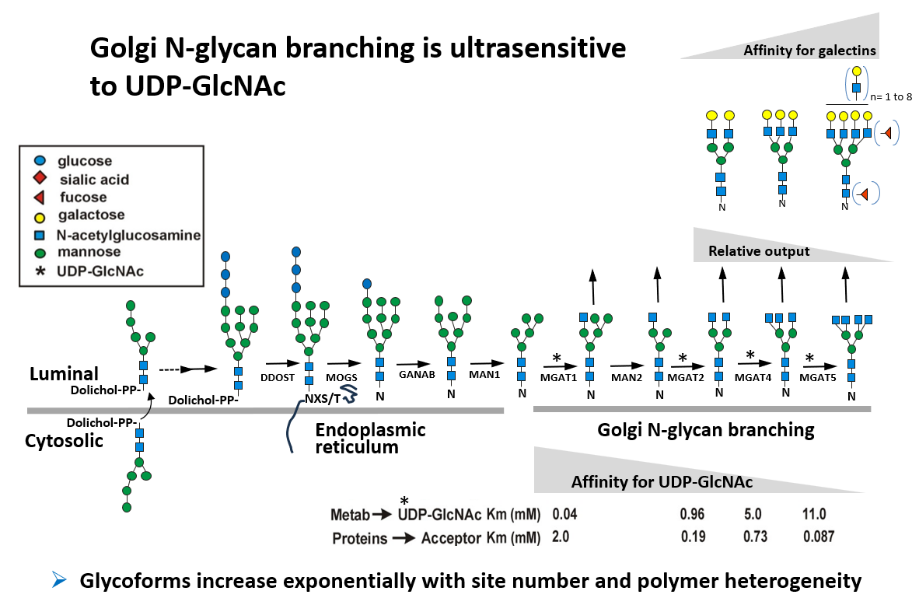 These enzyme reactions require uridine diphosphate N-acetylglucosamine (UDP-GlcNAc), generated by the hexosamine biosynthesis pathway (HBP) from glucose, glutamine and acetyl-coenzyme A. The GlcNAc-branched N-glycans are elongated by additional enzymes creating β-galactoside binding sites for Galectins, which crosslink and alter the dynamics of glycoproteins at the cell surface (
These enzyme reactions require uridine diphosphate N-acetylglucosamine (UDP-GlcNAc), generated by the hexosamine biosynthesis pathway (HBP) from glucose, glutamine and acetyl-coenzyme A. The GlcNAc-branched N-glycans are elongated by additional enzymes creating β-galactoside binding sites for Galectins, which crosslink and alter the dynamics of glycoproteins at the cell surface (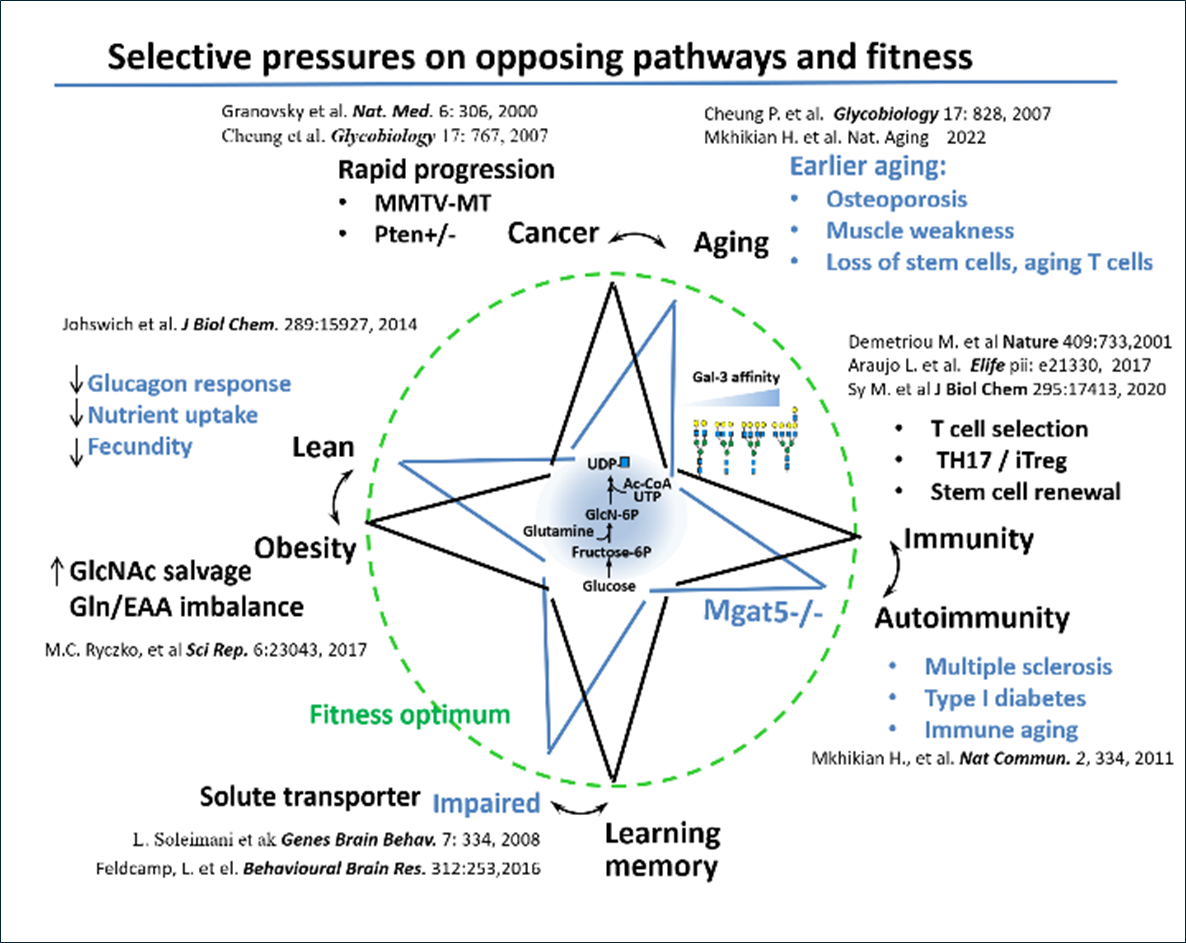 The Galectin lattice regulates opposing pathways including T cell receptor and CTLA-4 in lymphocytes (
The Galectin lattice regulates opposing pathways including T cell receptor and CTLA-4 in lymphocytes (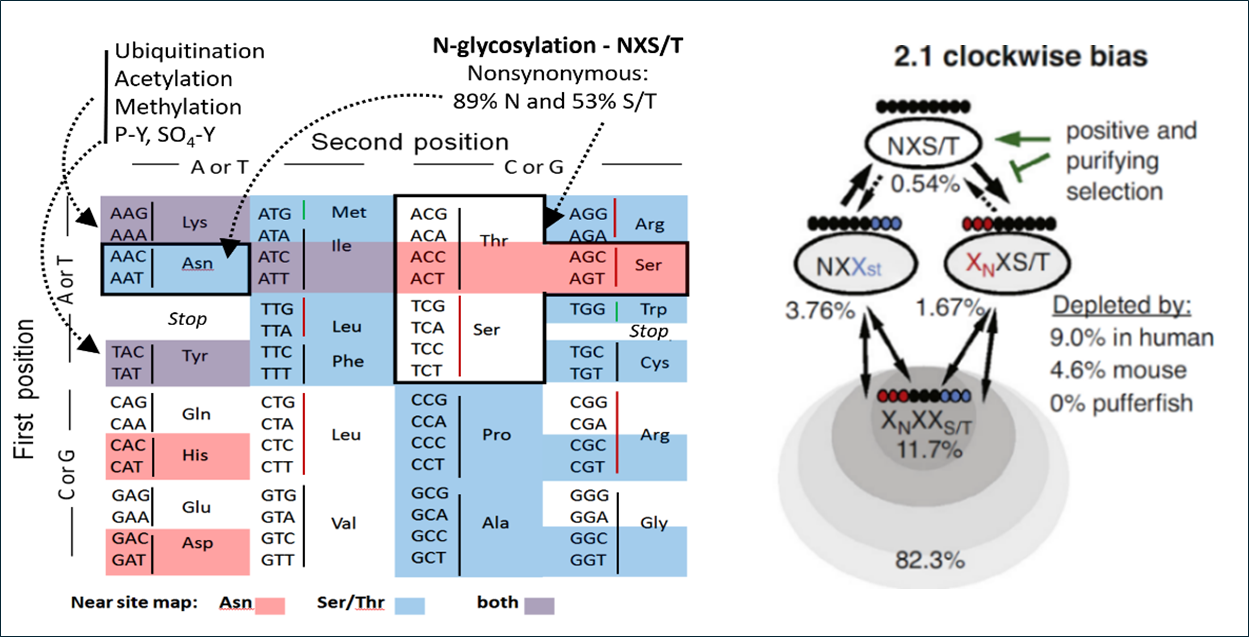 intermediates leading to the universal code ultimately minimizing the cost of information flow while maximizing amino acid diversity. However, mapping up to 25 amino acids in the triplet nucleotide format is possible based on information theory. Thus, fixation of the genetic code appears to have compromised on the total number of amino acids to allow for greater asymmetries in codon number per amino acid. Our phylogenetic analysis suggests that this asymmetry increases mutational experimentation with amino acids that are more likely to yield phenotypic diversity. Lys, and Asn are each encoded by two A-rich codons and therefore more likely to undergo nonsynonymous mutation than amino with more codons. Site loss followed by site gain is more likely to occur elsewhere, thus enhanced rate of experimentation with PTM sites (methylation, acetylation, ubiquitination and N-glycosylation). N-glycosylation sites are ~98% at bipartite motif NXS/T(X≠ P) by encoded (i.e., two Asn and ten Ser/Thr codons), a consistency and strong asymmetry that allowed us to measure repositioning of sites and the effect of coding asymmetry. The data is consistent with a clockwise loss-gain cycle that has repositioned sites with increasing positive selection during metazoan evolution (
intermediates leading to the universal code ultimately minimizing the cost of information flow while maximizing amino acid diversity. However, mapping up to 25 amino acids in the triplet nucleotide format is possible based on information theory. Thus, fixation of the genetic code appears to have compromised on the total number of amino acids to allow for greater asymmetries in codon number per amino acid. Our phylogenetic analysis suggests that this asymmetry increases mutational experimentation with amino acids that are more likely to yield phenotypic diversity. Lys, and Asn are each encoded by two A-rich codons and therefore more likely to undergo nonsynonymous mutation than amino with more codons. Site loss followed by site gain is more likely to occur elsewhere, thus enhanced rate of experimentation with PTM sites (methylation, acetylation, ubiquitination and N-glycosylation). N-glycosylation sites are ~98% at bipartite motif NXS/T(X≠ P) by encoded (i.e., two Asn and ten Ser/Thr codons), a consistency and strong asymmetry that allowed us to measure repositioning of sites and the effect of coding asymmetry. The data is consistent with a clockwise loss-gain cycle that has repositioned sites with increasing positive selection during metazoan evolution (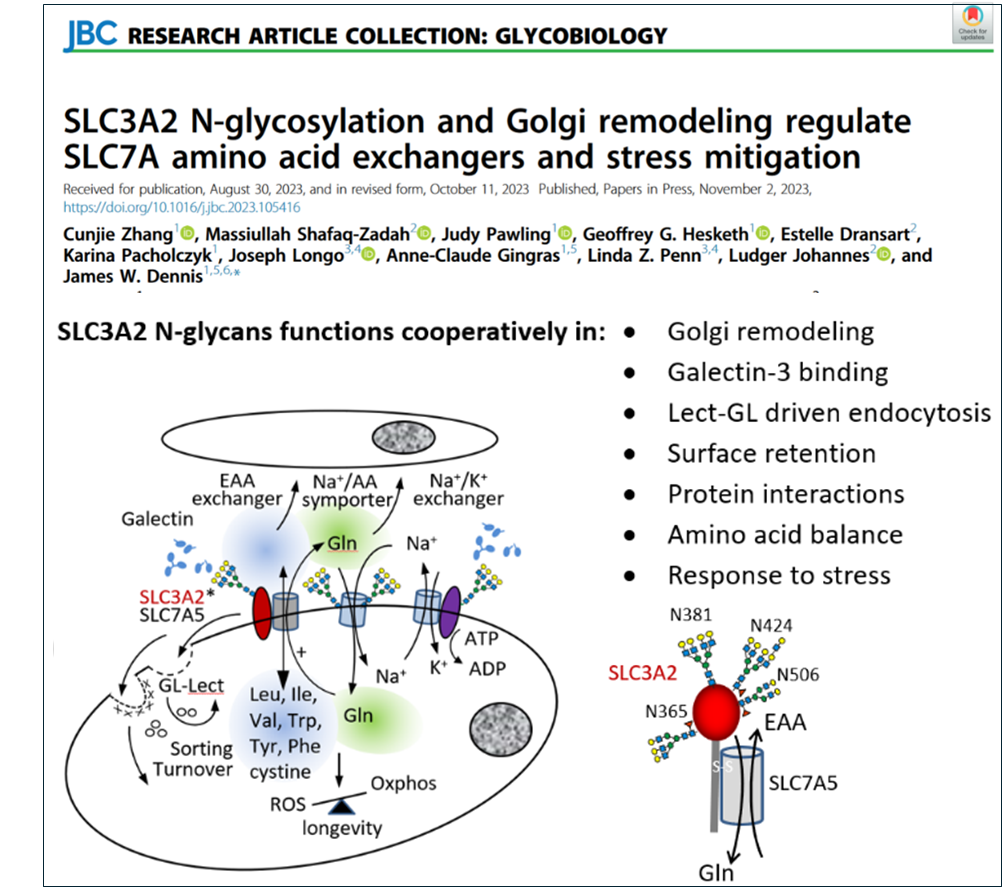 The import and balance of essential amino acid (EAA) in cells is dependent on nutrient flux to HBP and N-glycan branching on SLC3A2 (4F2hc, CD98), a transmembrane glycoprotein that forms a heterodimer with SLC7A family of essential amino acid exchangers. SLC3A2 bringing N-glycans to the heterodimer, as these exchangers are not N-glycosylated. Selective pressures on SLC3A2 N-glycosylation sites have been greater than other adaptors to ion transporters. Ancestral sites at four of eight positions in rodents varied locally by about 3-6 AAs over mammalian evolution but are absent in primates where a novel site was acquired at N381, and the other three sites at N365, N424 and N506 are conserved. We identify unique N-glycan structural profiles at each of the four sites and show that residue N381 and N365 of SLC3A2 interact with the Galectin-3 lattice and promote SLC3A2*SLC7A5 surface retention, transport activity and clustering with amino acid/Na+ symporters (SLC1A4, SLC1A5), and regulate sorting turnover by GL-Lect. EAA in SLC3A2 KO cells were decreased, while HBP substrates and UDP-GlcNAc increased, suggesting positive feedback directed at rebalancing EAA/Gln/Glu by increasing N-glycan branching (
The import and balance of essential amino acid (EAA) in cells is dependent on nutrient flux to HBP and N-glycan branching on SLC3A2 (4F2hc, CD98), a transmembrane glycoprotein that forms a heterodimer with SLC7A family of essential amino acid exchangers. SLC3A2 bringing N-glycans to the heterodimer, as these exchangers are not N-glycosylated. Selective pressures on SLC3A2 N-glycosylation sites have been greater than other adaptors to ion transporters. Ancestral sites at four of eight positions in rodents varied locally by about 3-6 AAs over mammalian evolution but are absent in primates where a novel site was acquired at N381, and the other three sites at N365, N424 and N506 are conserved. We identify unique N-glycan structural profiles at each of the four sites and show that residue N381 and N365 of SLC3A2 interact with the Galectin-3 lattice and promote SLC3A2*SLC7A5 surface retention, transport activity and clustering with amino acid/Na+ symporters (SLC1A4, SLC1A5), and regulate sorting turnover by GL-Lect. EAA in SLC3A2 KO cells were decreased, while HBP substrates and UDP-GlcNAc increased, suggesting positive feedback directed at rebalancing EAA/Gln/Glu by increasing N-glycan branching (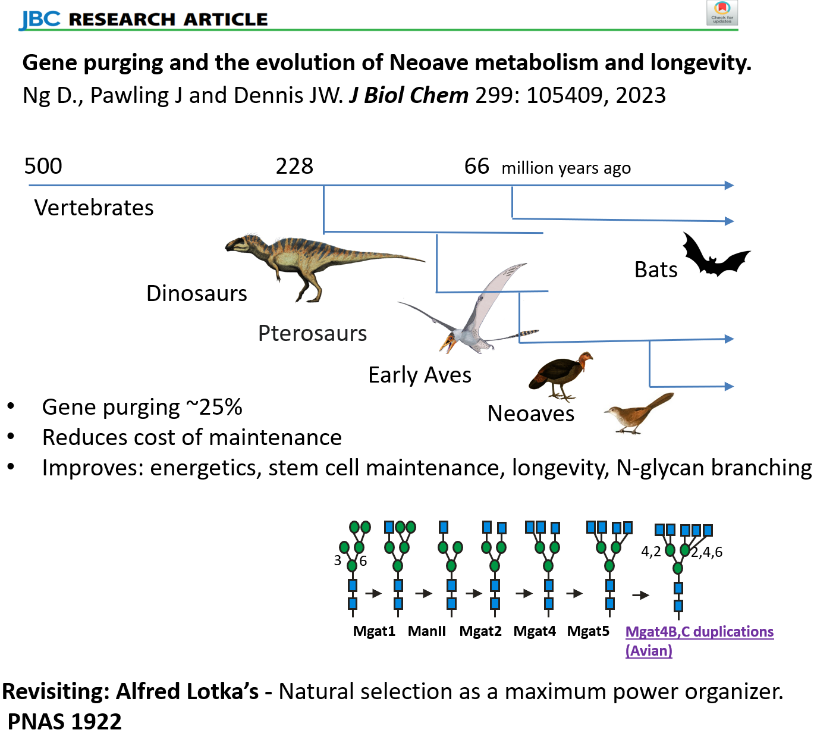 Intriguingly, the evolution of modern birds (Neoaves) has entailed the purging of genes (~25% coding sequences) including SLC3A2, SLC7A5, -7, -8, -10, and SLC1A4, -5, largely removing BCAA exchangers and their interacting Na(+)/Gln symporters in pursuit of improved energetics. Notably, export of Gln with the import of EAA by SLC3A2*SLC7A5 requires the recovery of Gln by AA/Na+ symporter at the cost (ATP) of maintaining the Na+/K+ gradient by the ATPase exchangers. Duplications of the bidirectional α-ketoacid transporters SLC16A3, SLC16A7, the cystine transporters SLC7A9, SLC7A11, and N-glycan branching enzymes MGAT4B, MGAT4C in Neoaves suggests a shift to the transport of deaminated essential amino acid, and stronger mitigation of oxidative stress supported by the Galectin lattice. We suggest that Alfred Lotka's theory of natural selection as a maximum power organizer (PNAS 8:151,1922) made an unusually large contribution to Neoave evolution.
Intriguingly, the evolution of modern birds (Neoaves) has entailed the purging of genes (~25% coding sequences) including SLC3A2, SLC7A5, -7, -8, -10, and SLC1A4, -5, largely removing BCAA exchangers and their interacting Na(+)/Gln symporters in pursuit of improved energetics. Notably, export of Gln with the import of EAA by SLC3A2*SLC7A5 requires the recovery of Gln by AA/Na+ symporter at the cost (ATP) of maintaining the Na+/K+ gradient by the ATPase exchangers. Duplications of the bidirectional α-ketoacid transporters SLC16A3, SLC16A7, the cystine transporters SLC7A9, SLC7A11, and N-glycan branching enzymes MGAT4B, MGAT4C in Neoaves suggests a shift to the transport of deaminated essential amino acid, and stronger mitigation of oxidative stress supported by the Galectin lattice. We suggest that Alfred Lotka's theory of natural selection as a maximum power organizer (PNAS 8:151,1922) made an unusually large contribution to Neoave evolution.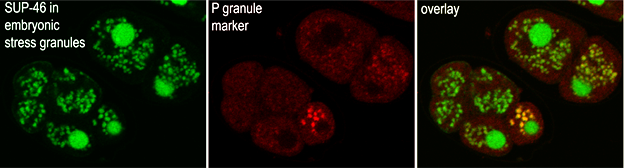 HNRNPM and MYEF2 are human homologs of SUP-46 also found in RNA stress granules. SUP-46 regulated a set of transcripts that overlap extensively with those regulated by the small RNA-binding Argonaute proteins, required for gametogenesis, early development and stress mitigation. Importantly, SUP-46 has an essential function in preserving paternally mediated transgenerational germline immortality.
HNRNPM and MYEF2 are human homologs of SUP-46 also found in RNA stress granules. SUP-46 regulated a set of transcripts that overlap extensively with those regulated by the small RNA-binding Argonaute proteins, required for gametogenesis, early development and stress mitigation. Importantly, SUP-46 has an essential function in preserving paternally mediated transgenerational germline immortality. 